High Protein Diet Induces Oxidative Stress in Rat Cerebral Cortex and Hypothalamus

 ,
,  ,
,  and
and
Abstract
1. Introduction
2. Results
2.1. Effects of High Protein Diet on Body Weight and Plasma Metabolic Parameters
2.2. Non-Enzymatic and Enzymatic Antioxidants, Total Antioxidant/Oxidant Status, and Oxidative Damage Product Level in Plasma/Serum
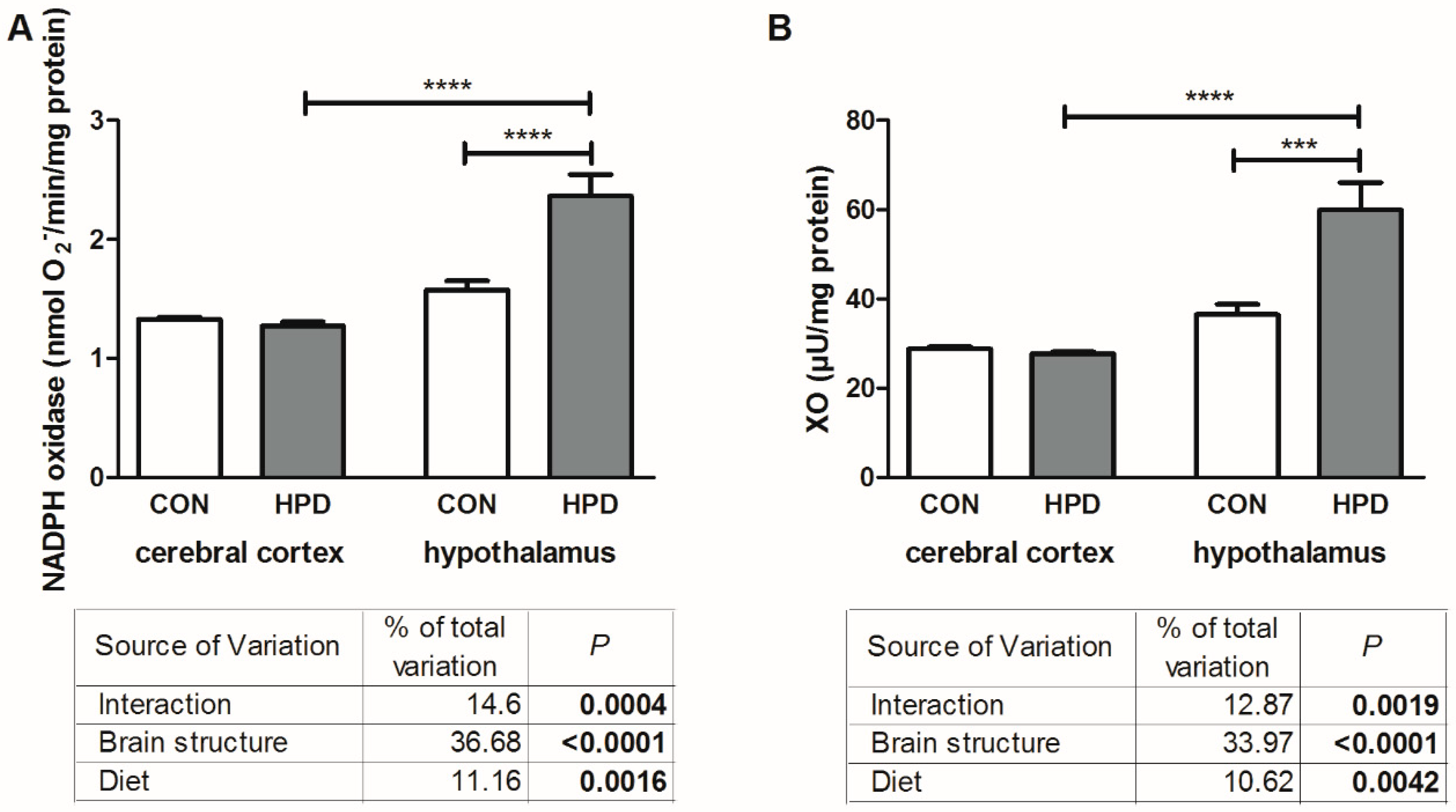
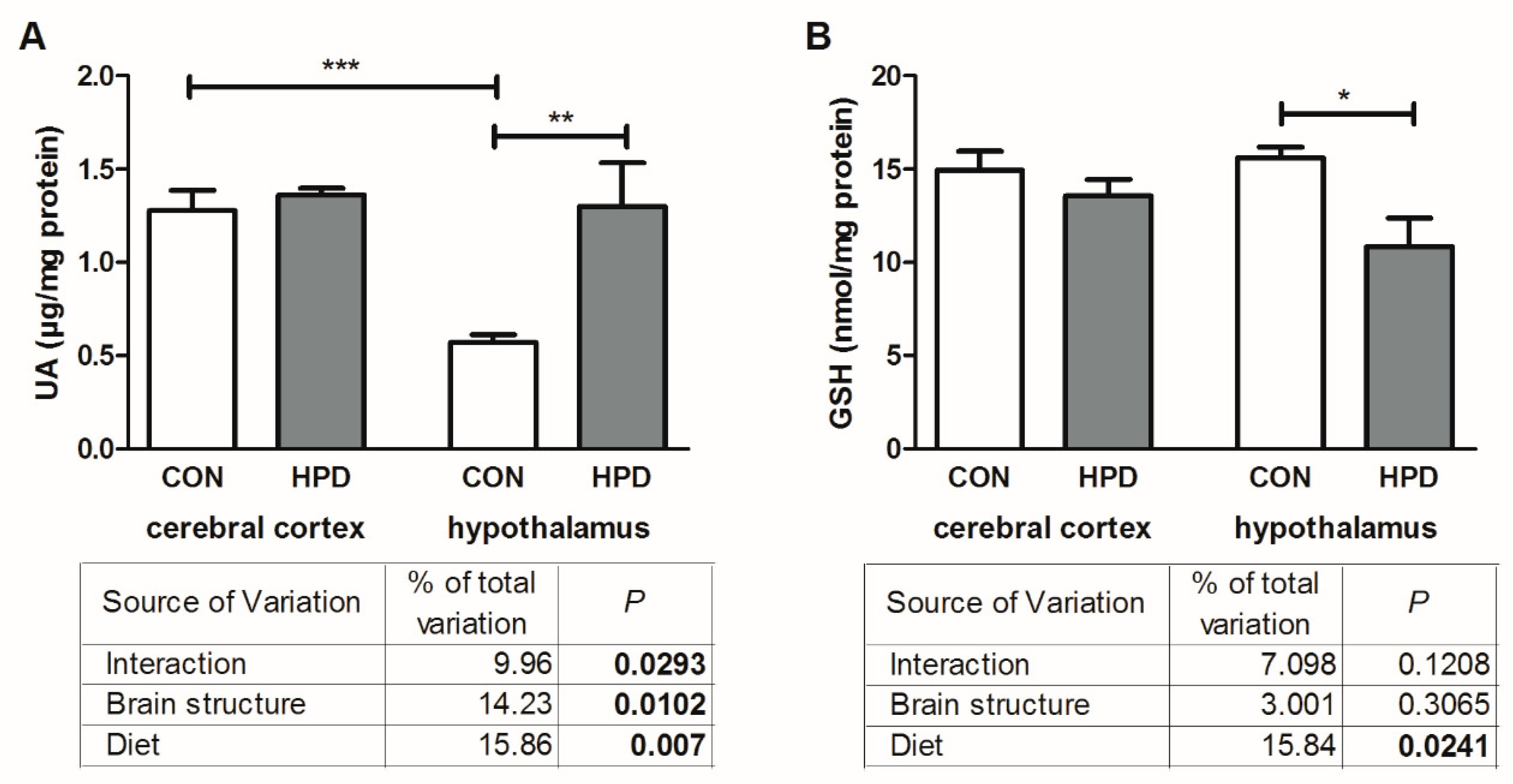
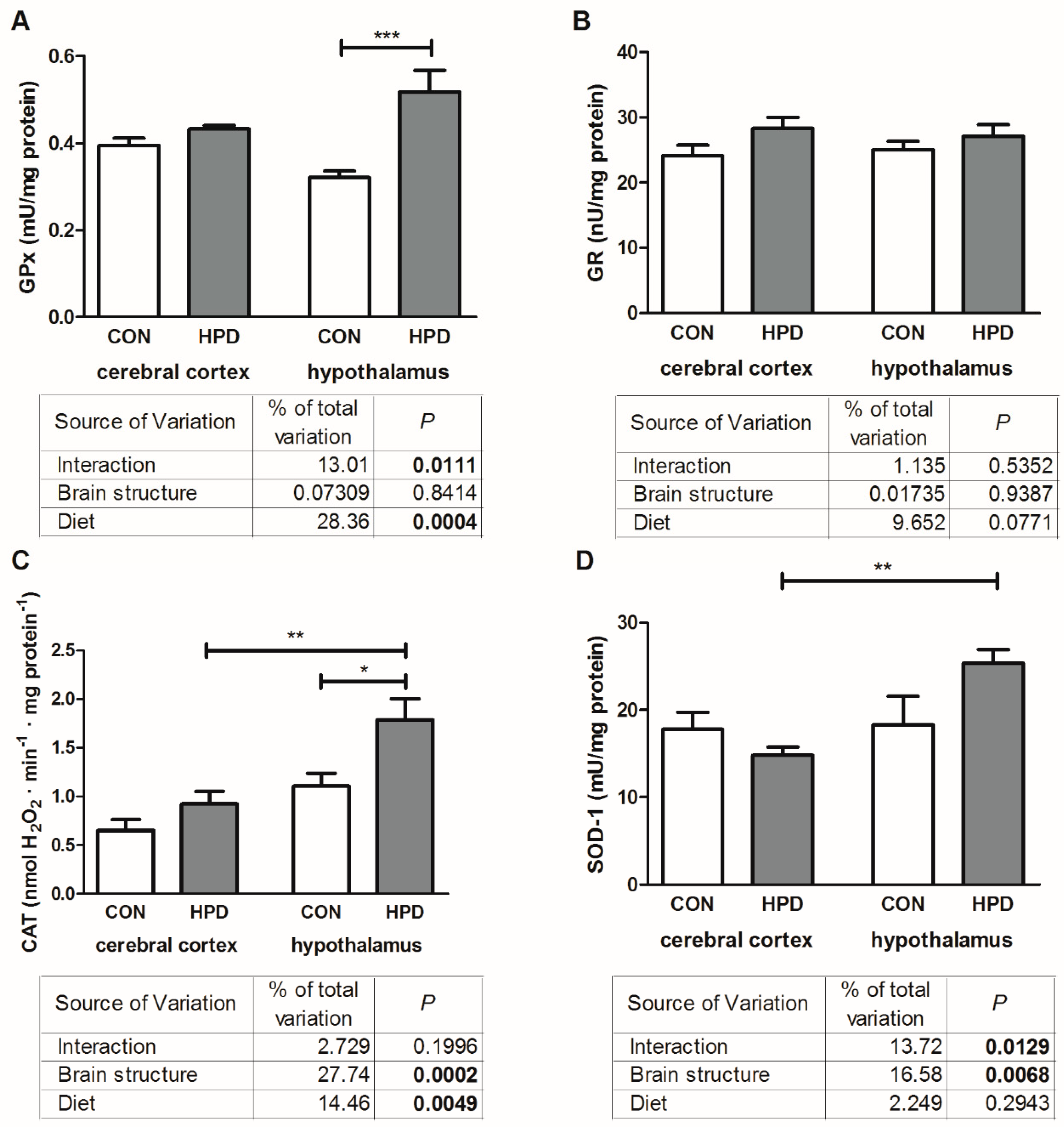
2.3. Pro-Oxidant Enzymes and Antioxidants in Cerebral Cortex and Hypothalamus
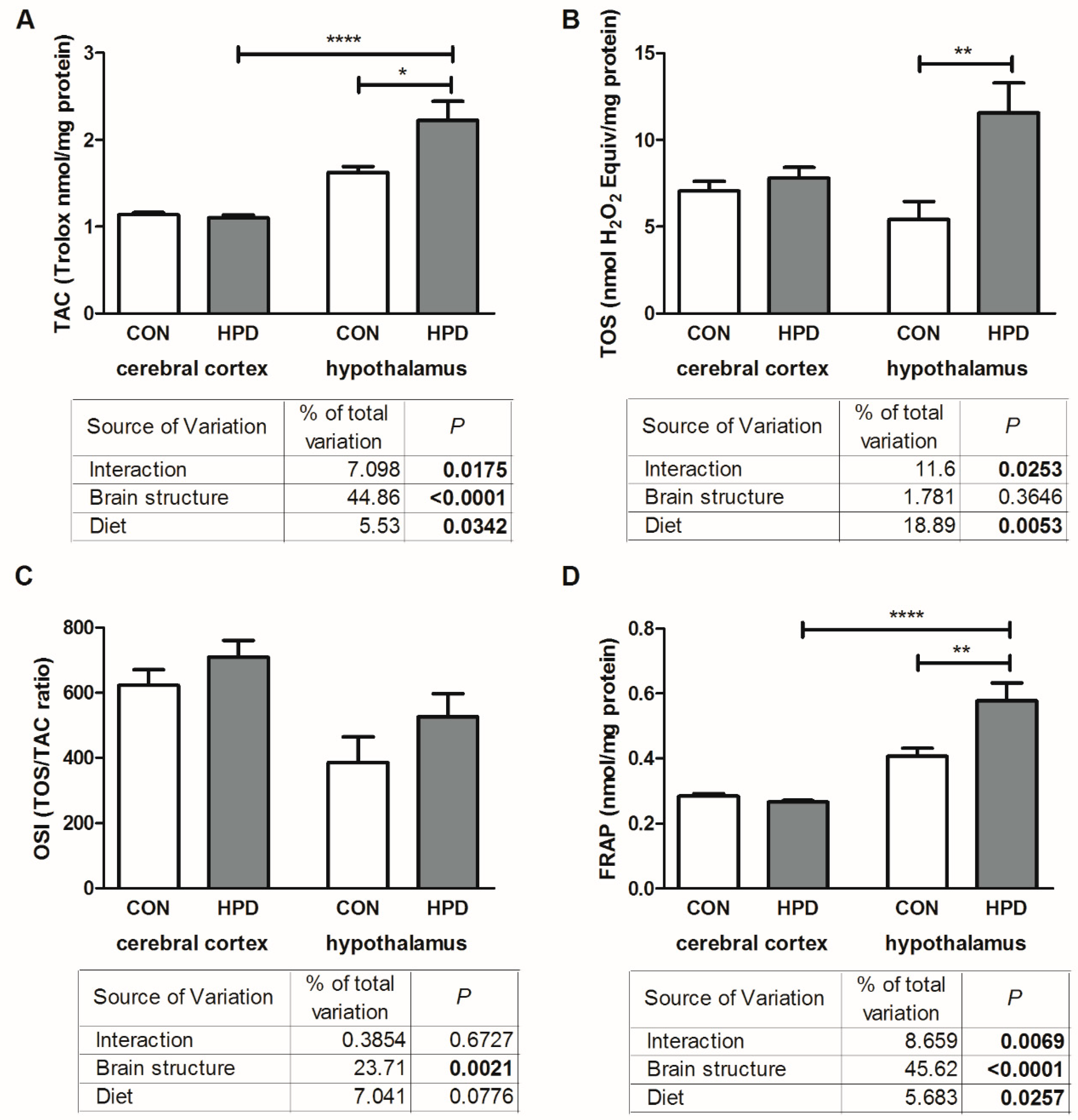
2.4. Total Antioxidant/Oxidant Status in Cerebral Cortex and Hypothalamus
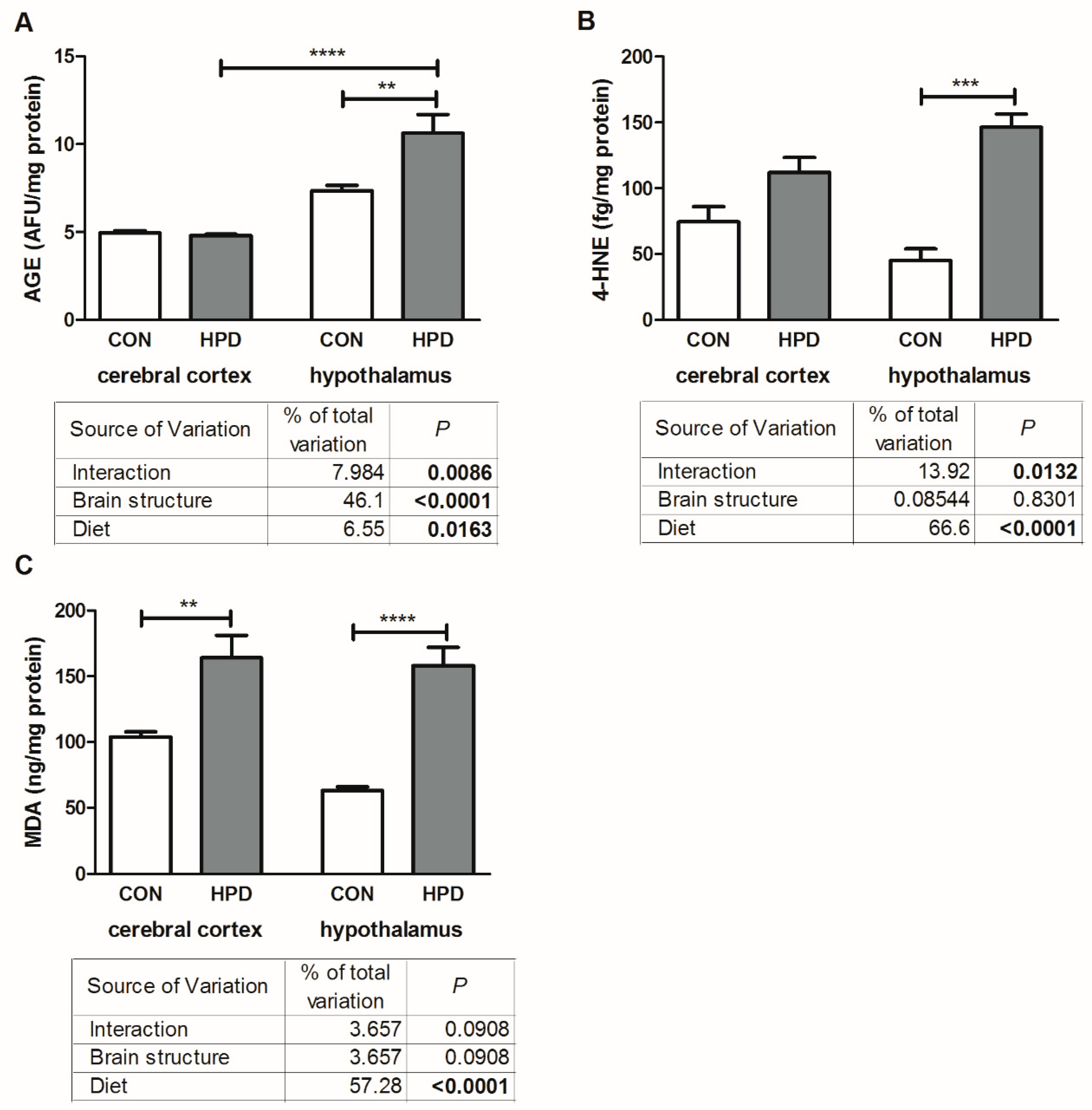
2.5. Oxidative Damage Products in Cerebral Cortex and Hypothalamus
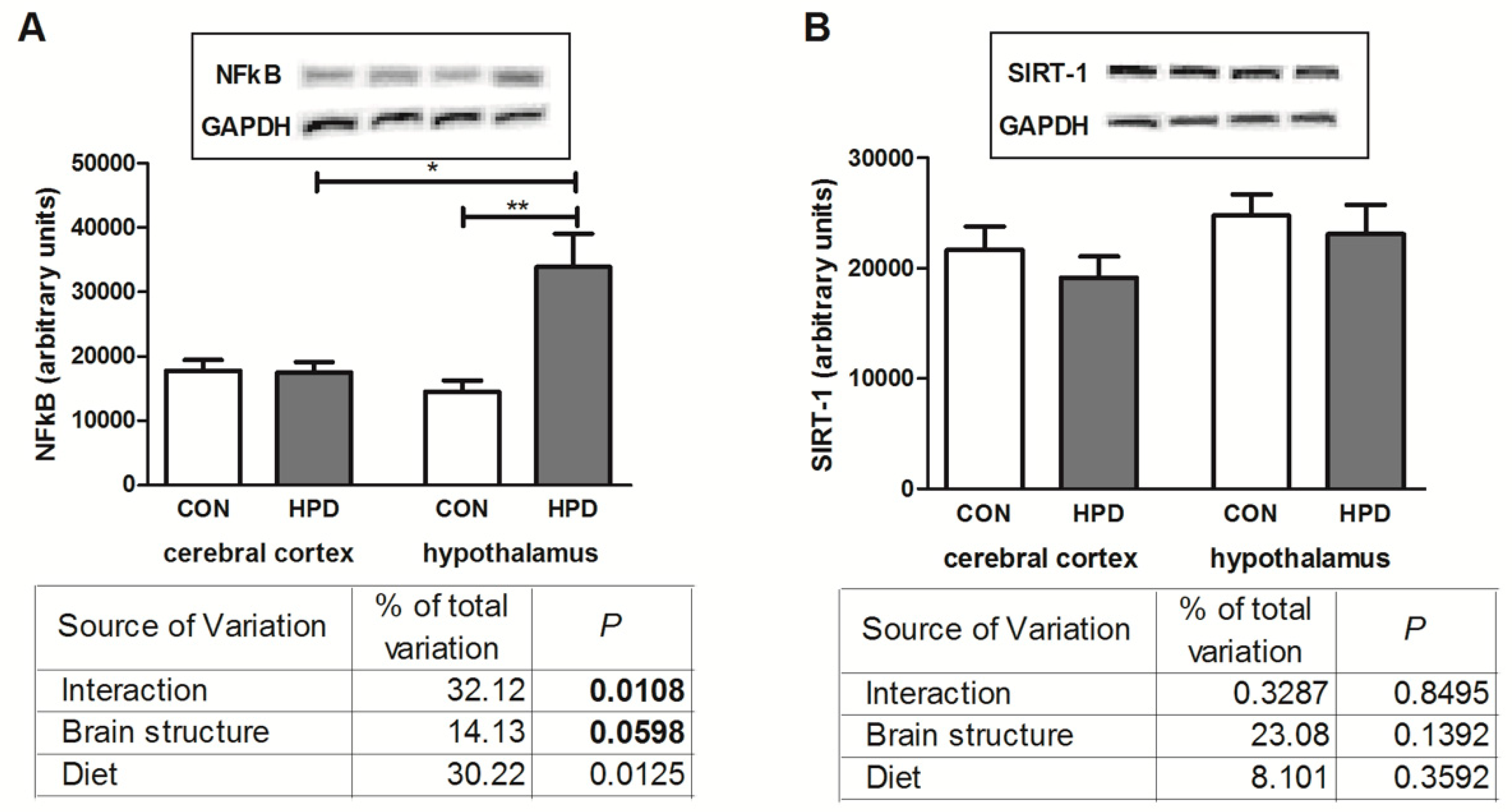
2.6. NFκB Expression and SIRT-1
3. Discussion
4. Materials and Methods
4.1. Plasma Insulin, Glucose, Adiponectin, and Leptin Concentrations
4.2. Pro-Oxidant Enzymes and Antioxidants in Cerebral Cortex and Hypothalamus
4.3. Total Antioxidant/Oxidant Status
4.4. Oxidative Modification Products
4.5. Western Blot Analysis
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 4-HNE | 4-hydroxynonenal |
| AGE | advanced glycation end products |
| BBB | blood–brain barrier |
| BCA | bicinchoninic acid |
| BCAA | branched-chain amino acids |
| BHT | butylated hydroxytoluene |
| CAT | catalase |
| CNS | central nervous system |
| FRAP | ferric reducing ability of plasma |
| GAPDH | glyceraldehyde 3-phosphate dehydrogenase |
| GPx | glutathione peroxidase |
| GR | glutathione reductase |
| GSH | reduced glutathione |
| HOMA-IR | homeostatic model assessment of β-cell function and insulin resistance |
| HPD | high protein diet |
| MDA | malondialdehyde |
| NADPH | nicotinamide adenine dinucleotide phosphate |
| NFκB | nuclear factor-κB |
| OS | oxidative stress |
| OSI | oxidative stress index |
| PBS | phosphate buffered saline |
| PUFAs | polyunsaturated fatty acids |
| RAGE | receptor for advanced glycation end products |
| ROS | reactive oxygen species |
| SDS-PAGE | sodium dodecyl sulfate-polyacrylamide gel electrophoresis |
| SIRT-1 | sirtuin 1 |
| SOD-1 | superoxide dismutase-1 |
| TAC | total antioxidant capacity |
| TBARS | thiobarbituric acid reactive substances |
| TNF-alpha | tumor necrosis factor alpha |
| TOS | total oxidant status |
| UA | uric acid |
| WHO | World Health Organization |
References
- Keller, U. Dietary proteins in obesity and in diabetes. Int. J. Vitam. Nutr. Res. 2011, 81, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Astrup, A.; Raben, A.; Geiker, N. The role of higher protein diets in weight control and obesity-related comorbidities. Int. J. Obes. 2015, 39, 721–726. [Google Scholar] [CrossRef] [PubMed]
- Halton, T.L.; Hu, F.B. The effects of high protein diets on thermogenesis, satiety and weight loss: A critical review. J. Am. Coll. Nutr. 2004, 23, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Leidy, H.J.; Clifton, P.M.; Astrup, A.; Wycherley, T.P.; Westerterp-Plantenga, M.S.; Luscombe-Marsh, N.D.; Woods, S.C.; Mattes, R.D. The role of protein in weight loss and maintenance. Am. J. Clin. Nutr. 2015, 101, 1320–1329. [Google Scholar] [CrossRef] [PubMed]
- WHO. Energy and Protein Requirements: Report of a Joint FAO/WHO/UNU Expert Consultation (724); WHO: Geneva, Switzerland, 1985. [Google Scholar]
- Metges, C.C.; Barth, C.A. Metabolic consequences of a high dietary-protein intake in adulthood: Assessment of the available evidence. J. Nutr. 2000, 130, 886–889. [Google Scholar] [CrossRef]
- Delimaris, I. Adverse effects associated with protein intake above the recommended dietary allowance for adults. ISRN Nutr. 2013, 2013, 126929. [Google Scholar] [CrossRef] [PubMed]
- Newgard, C.B. Interplay between lipids and branched-chain amino acids in development of insulin resistance. Cell Metab. 2012, 15, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Petzke, K.J.; Proll, J.; Brückner, J.; Metges, C.C. Plasma protein carbonyl concentration is not enhanced by chronic intake of high-protein diets in adult rats. J. Nutr. Biochem. 1999, 10, 268–273. [Google Scholar] [CrossRef]
- Gu, C.; Shi, Y.; Le, G. Effect of dietary protein level and origin on the redox status in the digestive tract of mice. Int. J. Mol. Sci. 2008, 9, 464–475. [Google Scholar] [CrossRef]
- Gu, C.; Xu, H. Effect of oxidative damage due to excessive protein ingestion on pancreas function in mice. Int. J. Mol. Sci. 2010, 11, 4591–4600. [Google Scholar] [CrossRef]
- Kołodziej, U.; Maciejczyk, M.; Niklińska, W.; Waszkiel, D.; Żendzian-Piotrowska, M.; Żukowski, P.; Zalewska, A. Chronic high-protein diet induces oxidative stress and alters the salivary gland function in rats. Arch. Oral Biol. 2017, 84, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Journel, M.; Chaumontet, C.; Darcel, N.; Fromentin, G.; Tomé, D. Brain responses to high-protein diets. Adv. Nutr. 2012, 3, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Lovekamp-Swan, T.; Glendenning, M.L.; Schreihofer, D.A. A high soy diet enhances neurotropin receptor and Bcl-XLgene expression in the brains of ovariectomized female rats. Brain Res. 2007, 1159, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Méndez-López, M.; Méndez, M.; Arias, J.; Arias, J.L. Effects of a high protein diet on cognition and brain metabolism in cirrhotic rats. Physiol. Behav. 2015, 149, 220–228. [Google Scholar] [CrossRef]
- Pedrini, S.; Thomas, C.; Brautigam, H.; Schmeidler, J.; Ho, L.; Fraser, P.; Westaway, D.; Hyslop, P.S.G.; Martins, R.N.; Buxbaum, J.D.; et al. Dietary composition modulates brain mass and solubilizable A levels in a mouse model of aggressive Alzheimer’s amyloid pathology. Mol. Neurodegener. 2009, 4. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Michaelis, E.K. Selective neuronal vulnerability to oxidative stress in the brain. Front. Aging Neurosci. 2010, 2, 12. [Google Scholar] [CrossRef]
- Camiletti-Moirón, D.; Aparicio, V.A.; Aranda, P.; Radak, Z. Does exercise reduce brain oxidative stress? A systematic review. Scand. J. Med. Sci. Sport. 2013, 23, 202–212. [Google Scholar] [CrossRef]
- Chiurchiù, V.; Orlacchio, A.; Maccarrone, M. Is modulation of oxidative stress an answer? the state of the art of redox therapeutic actions in neurodegenerative diseases. Oxid. Med. Cell. Longev. 2016, 2016, 1–11. [Google Scholar] [CrossRef]
- Maciejczyk, M.; Żebrowska, E.; Zalewska, A.; Chabowski, A. Redox Balance, Antioxidant Defense, and Oxidative Damage in the Hypothalamus and Cerebral Cortex of Rats with High Fat Diet-Induced Insulin Resistance. Oxid. Med. Cell. Longev. 2018, 2018. [Google Scholar] [CrossRef]
- Mandal, P.K.; Tripathi, M.; Sugunan, S. Brain oxidative stress: Detection and mapping of anti-oxidant marker “Glutathione” in different brain regions of healthy male/female, MCI and Alzheimer patients using non-invasive magnetic resonance spectroscopy. Biochem. Biophys. Res. Commun. 2012, 417, 43–48. [Google Scholar] [CrossRef]
- Choromańska, M.; Klimiuk, A.; Kostecka-Sochoń, P.; Wilczyńska, K.; Kwiatkowski, M.; Okuniewska, N.; Waszkiewicz, N.; Zalewska, A.; Maciejczyk, M. Antioxidant defence, oxidative stress and oxidative damage in saliva, plasma and erythrocytes of dementia patients. Can salivary AGE be a marker of dementia? Int. J. Mol. Sci. 2017, 18, 2205. [Google Scholar] [CrossRef] [PubMed]
- Maciejczyk, M.; Mikoluc, B.; Pietrucha, B.; Heropolitanska-Pliszka, E.; Pac, M.; Motkowski, R.; Car, H. Oxidative stress, mitochondrial abnormalities and antioxidant defense in Ataxia-telangiectasia, Bloom syndrome and Nijmegen breakage syndrome. Redox Biol. 2017, 11, 375–383. [Google Scholar] [CrossRef]
- Falkowski, M.; Maciejczyk, M.; Koprowicz, T.; Mikołuć, B.; Milewska, A.; Zalewska, A.; Car, H. Whey Protein Concentrate WPC-80 Improves Antioxidant Defense Systems in the Salivary Glands of 14-Month Wistar Rats. Nutrients 2018, 10, 782. [Google Scholar] [CrossRef]
- Camiletti-Móiron, D.; Arianna Aparicio, V.; Nebot, E.; Medina, G.; Martínez, R.; Kapravelou, G.; Andrade, A.; Porres, J.M.; López-Jurado, M.; Aranda, P. High-protein diet induces oxidative stress in rat brain: Protective action of high-intensity exercise against lipid peroxidation. Nutr. Hosp. 2015, 31, 866–874. [Google Scholar] [CrossRef]
- Borys, J.; Maciejczyk, M.; Kretowski, A.J.; Antonowicz, B.; Ratajczak-Wrona, W.; Jablonska, E.; Zaleski, P.; Waszkiel, D.; Ladny, J.R.; Zukowski, P.; et al. The redox balance in erythrocytes, plasma, and periosteum of patients with titanium fixation of the jaw. Front. Physiol. 2017, 8, 1–11. [Google Scholar] [CrossRef]
- Lubitz, I.; Ricny, J.; Atrakchi-Baranes, D.; Shemesh, C.; Kravitz, E.; Liraz-Zaltsman, S.; Maksin-Matveev, A.; Cooper, I.; Leibowitz, A.; Uribarri, J.; et al. High dietary advanced glycation end products are associated with poorer spatial learning and accelerated Aβ deposition in an Alzheimer mouse model. Aging Cell 2016, 15, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Maciejczyk, M.; Żebrowska, E.; Chabowski, A. Insulin Resistance and Oxidative Stress in the Brain: What’s New? Int. J. Mol. Sci. 2019, 20, 874. [Google Scholar] [CrossRef]
- Uribarri, J.; Tuttle, K.R. Advanced glycation end products and nephrotoxicity of high-protein diets. Clin. J. Am. Soc. Nephrol. 2006, 1, 1293–1299. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef]
- De Assis, D.R.; Maria, R.C.; Ferreira, G.C.; Schuck, P.F.; Latini, A.; Dutra-Filho, C.S.; Wannmacher, C.M.D.; Wyse, A.T.S.; Wajner, M. Na+,K+ ATPase activity is markedly reduced by cis-decenoic acid in synaptic plasma membranes from cerebral cortex of rats. Exp. Neurol. 2005, 197, 143–149. [Google Scholar] [CrossRef]
- Klaunig, J.E.; Wang, Z.; Pu, X.; Zhou, S. Oxidative stress and oxidative damage in chemical carcinogenesis. Toxicol. Appl. Pharmacol. 2011, 254, 86–99. [Google Scholar] [CrossRef] [PubMed]
- Enciu, A.M.; Gherghiceanu, M.; Popescu, B.O. Triggers and effectors of oxidative stress at blood-brain barrier level: Relevance for brain ageing and neurodegeneration. Oxid. Med. Cell. Longev. 2013, 2013, 297512. [Google Scholar] [CrossRef] [PubMed]
- Snelson, M.; Mamo, J.C.L.; Lam, V.; Giles, C.; Takechi, R. Differential effects of high-protein diets derived from soy and casein on blood–brain barrier integrity in mild-type mice. Front. Nutr. 2017, 4, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.J.; Yamada, K.; Sugisawa, A.; Saito, K.; Miyajima, T.; Umegaki, K. Enhanced oxidative damage induced by total body irradiation in mice fed a low protein diet. Indian J. Radiat. Biol. 2002, 78, 425–432. [Google Scholar] [CrossRef]
- Shin, S.J. Does a high protein diet induce oxidative damage? SM J. Food Nutr. Disord. 2015, 1, 1001. [Google Scholar]
- Soulsby, M.E.; Phillips, B.; Chowdhury, P. Effects of soy-protein diet on elevated brain lipid peroxide levels induced by simulated weightlessness. Ann. Clin. Lab. Sci. 2004, 34, 103–106. [Google Scholar]
- Jean, C.; Rome, S.; Aattouri, N.; Fromentin, G.; Achagiotis, C.L.; Tome, D. Metabolic evidence for adaptation to a high protein diet in rats. J. Nutr. 2001, 131, 91–98. [Google Scholar] [CrossRef]
- Petzke, K.J.; Elsner, A.; Proll, J.; Thielecke, F.; Metges, C.C. Long-term high protein intake does not increase oxidative stress in rats. J. Nutr. 2000, 130, 2889–2896. [Google Scholar] [CrossRef]
- Villa, R.F.; Ferrari, F.; Gorini, A. Energy metabolism of rat cerebral cortex, hypothalamus and hypophysis during ageing. Neuroscience 2012, 227, 55–66. [Google Scholar] [CrossRef]
- Morrison, C.D.; Pistell, P.J.; Ingram, D.K.; Johnson, W.D.; Liu, Y.; Fernandez-Kim, S.O.; White, C.L.; Purpera, M.N.; Uranga, R.M.; Bruce-Keller, A.J.; et al. High fat diet increases hippocampal oxidative stress and cognitive impairment in aged mice: Implications for decreased Nrf2 signaling. J. Neurochem. 2010, 114, 1581–1589. [Google Scholar] [CrossRef]
- Borys, J.; Maciejczyk, M.; Antonowicz, B.; Krętowski, A.; Waszkiel, D.; Bortnik, P.; Czarniecka-Bargłowska, K.; Kocisz, M.; Szulimowska, J.; Czajkowski, M.; et al. Exposure to Ti4Al4V Titanium Alloy Leads to Redox Abnormalities, Oxidative Stress, and Oxidative Damage in Patients Treated for Mandible Fractures. Oxid. Med. Cell. Longev. 2018, 2018, 1–10. [Google Scholar] [CrossRef]
- Żukowski, P.; Maciejczyk, M.; Matczuk, J.; Kurek, K.; Waszkiel, D.; Żendzian-Piotrowska, M.; Zalewska, A. Effect of N-acetylcysteine on antioxidant defense, oxidative modification, and salivary gland function in a rat model of insulin resistance. Oxid. Med. Cell. Longev. 2018, 2018, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Griendling, K.K.; Minieri, C.A.; Ollerenshaw, J.D.; Alexander, R.W. Angiotensin II stimulates NADH and NADPH oxidase activity in cultured vascular smooth muscle cells. Circ. Res. 1994, 74, 1141–1148. [Google Scholar] [CrossRef]
- Prajda, N.; Weber, G. Malignant transformation-linked imbalance: Decreased xanthine oxidase activity in hepatomas. FEBS Lett. 1975, 59, 245–249. [Google Scholar] [CrossRef]
- Paglia, D.E.; Valentine, W.N. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 1967, 70, 158–169. [Google Scholar] [PubMed]
- Mize, C.E.; Langdon, R.G. Hepatic glutathione reductase. I. Purification and general kinetic properties. J. Biol. Chem. 1962, 237, 1589–1595. [Google Scholar]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1984; Volume 105, pp. 121–126. ISBN 9780121820053. [Google Scholar]
- Misra, H.P.; Fridovich, I. The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [PubMed]
- Moron, M.S.; Depierre, J.W.; Mannervik, B. Levels of glutathione, glutathione reductase and glutathione S-transferase activities in rat lung and liver. Biochim. Biophys. Acta - Gen. Subj. 1979, 582, 67–78. [Google Scholar] [CrossRef]
- Walker, J.M. The bicinchoninic acid (BCA) assay for protein quantitation. In Basic Protein and Peptide Protocols; Humana Press: Totowa, NJ, USA, 1994; Volume 32, pp. 5–8. [Google Scholar]
- Erel, O. A novel automated direct measurement method for total antioxidant capacity using a new generation, more stable ABTS radical cation. Clin. Biochem. 2004, 37, 277–285. [Google Scholar] [CrossRef]
- Erel, O. A new automated colorimetric method for measuring total oxidant status. Clin. Biochem. 2005, 38, 1103–1111. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Methods Enzymol. 1978, 52, 302–310. [Google Scholar] [PubMed]
- Mikłosz, A.; Łukaszuk, B.; Żendzian-Piotrowska, M.; Brańska-Januszewska, J.; Ostrowska, H.; Chabowski, A. Challenging of AS160/TBC1D4 Alters Intracellular Lipid milieu in L6 Myotubes Incubated with Palmitate. J. Cell. Physiol. 2017, 232, 2373–2386. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | CON | HPD |
|---|---|---|
| Body weight (g) | 341 ± 2.49 | 350 ± 7.57 |
| Glucose concentration (mg/dL) | 99.8 ± 2.49 | 101 ± 5.12 |
| Insulin concentration (µU/mL) | 4.75 ± 0.02 | 4.95 ± 0.14 |
| HOMA-IR | 3.08 ± 0.17 | 3.56 ± 0.14 |
| Adiponectin (µg/mL) | 23.4 ± 0.57 | 22.4 ± 0.55 |
| Leptin (ng/mL) | 26.4 ± 0.63 | 24.8 ± 0.61 |
| Food intake (g/day) | 21.2 ± 0.84 | 16.2 ± 0.63 * |
| Energy intake (mJ/day) | 0.28 ± 0.01 | 0.27 ± 0.03 |
| Protein energy intake (mJ/day) | 0.06 ± 0.05 | 0.13 ± 0.04 * |
| Cerebral cortex total protein concentration (µg/mL) | 2742 ± 83.1 | 2796 ± 43.4 |
| Hypothalamus total protein concentration (µg/mL) | 1710 ± 66.1 | 1529 ± 53.3 |
| Parameter | CON | HPD |
|---|---|---|
| GPx (mU/mg protein) | 0.44 ± 0.02 | 0.64 ± 0.06 * |
| GR (nU/mg protein) | 11.8 ± 0.77 | 9.55 ± 0.32 |
| CAT (nmol H2O2·min−1·mg protein−1) | 6.02 ± 0.49 | 25.6 ± 1.52 * |
| SOD-1 (mU/mg protein) | 54.3 ± 2.02 | 42.8 ± 0.65 * |
| UA (µg/mg protein) | 2.71 ± 0.27 | 4.89 ± 0.58 * |
| GSH (nmol/mg protein) | 7.30 ± 0.71 | 3.06 ± 0.59 * |
| TAC (Trolox nmol/mg protein) | 3.94 ± 0.13 | 5.14 ± 0.51 |
| TOS (nmol H2O2 Equiv/mg protein) | 13.5 ± 2.87 | 27.5 ± 2.22 * |
| OSI (TOS/TAC ratio) | 344 ± 78.5 | 542 ± 38.9 |
| FRAP (nmol/mg protein) | 1.14 ± 0.09 | 1.51 ± 0.09 * |
| AGE (AFU/mg protein) | 3.26 ± 0.21 | 4.06 ± 0.21 * |
| 4-HNE (fg/mg protein) | 279 ± 62.6 | 1028 ± 85.7 * |
| MDA (ng/mg protein) | 471 ± 28.7 | 607 ± 44 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Żebrowska, E.; Maciejczyk, M.; Żendzian-Piotrowska, M.; Zalewska, A.; Chabowski, A. High Protein Diet Induces Oxidative Stress in Rat Cerebral Cortex and Hypothalamus. Int. J. Mol. Sci. 2019, 20, 1547. https://doi.org/10.3390/ijms20071547
Żebrowska E, Maciejczyk M, Żendzian-Piotrowska M, Zalewska A, Chabowski A. High Protein Diet Induces Oxidative Stress in Rat Cerebral Cortex and Hypothalamus. International Journal of Molecular Sciences. 2019; 20(7):1547. https://doi.org/10.3390/ijms20071547
Chicago/Turabian StyleŻebrowska, Ewa, Mateusz Maciejczyk, Małgorzata Żendzian-Piotrowska, Anna Zalewska, and Adrian Chabowski. 2019. "High Protein Diet Induces Oxidative Stress in Rat Cerebral Cortex and Hypothalamus" International Journal of Molecular Sciences 20, no. 7: 1547. https://doi.org/10.3390/ijms20071547
APA StyleŻebrowska, E., Maciejczyk, M., Żendzian-Piotrowska, M., Zalewska, A., & Chabowski, A. (2019). High Protein Diet Induces Oxidative Stress in Rat Cerebral Cortex and Hypothalamus. International Journal of Molecular Sciences, 20(7), 1547. https://doi.org/10.3390/ijms20071547